Boris Zimmermann and Achim Kohler
Department of Mathematical Sciences and Technology, Faculty of Environmental Science and Technology, Norwegian University of Life Sciences, Drøbakveien 31, 1432 Ås, Norway. E-mail: [email protected]
Introduction
Global climate change is already forcing plants to adapt, and it is foreseen that by the end of the century we can expect widespread disruption and extinction of species. Moreover, changes in plant genetic composition are expected as response to the selection pressure of climate change. However, the projections of future changes are limited by incomplete theoretical understanding and insufficient data to improve ecological models. Therefore, methodology that provides assessments of beneficial plant traits for the conditions which may prevail in the very near future is of utmost importance for estimating the impact of climate change on agronomy, forestry and the ecosystem in general. Innovative phenotyping and monitoring methods can also strengthen biotechnological development of new plant varieties, such as crops, that can maintain high yields under conditions that limit the productivity of current plants.
Seed plants (spermatophytes) comprise almost 90% of all plant species; they provide the foundation of human diets as well as timber and other material products. As with all land plants, they alternate between sporophyte generation that makes up almost their whole life cycle and gametophyte, a microscopic reproductive organism. Traditional plant phenotyping is focused on sporophytes, i.e. foliage, stems, roots and flowers. On the other hand, phenotyping of the male gametophyte, the pollen grain, is largely unexplored, given that characterisation is predominantly based on morphological features while biochemical properties are rarely considered.
Each pollen grain is a multicellular organism, the sole purpose of which is pollination: assisted transport by wind or animals from the parent plant to the reproductive organs of the complementary plant. Pollination results with the production of genetically diverse offspring, and is therefore a crucial step in the propagation of seed plants. Since the existence of a plant population vitally depends on mobility, survivability and the siring success of pollen, evolutionary imperatives have led to extraordinary diversification of the grains. As a result there are extreme differentiations among various morphological parameters, such as grain shape and size, number, type and position of apertures, and structure and sculpture of the grain wall (see, for example Figure 1).
 Branch of </em>Abies cephalonica<em> (Greek fir) during pollination; male cones (strobili) with yellow pollen powder are clearly visible. (B) Scanning electron microscope image of A</em>bies cephalonica<em> pollen grain in equatorial view; false colours depict saccus (upper region, two hemispherical substructures) and corpus (lower region, large hemispherical substructure) regions.")
Pollen identification provides useful information in various fields, for instance in public health for allergy forecasts, in ecology for monitoring of life cycles of vegetation, and in forensics for temporal and spatial determination of a criminal event. Unfortunately, current pollen studies are still confined to visual measurements of pollen morphologies under a microscope and the research has remained basically unchanged over the last hundred years. The identification requires laborious visual recognition of pollen grains from each plant taxon by a skilled microscopist with specific knowledge of pollen morphology. However, the last decade has seen a growing number of experimental works dedicated to an alternative and more objective pollen identification method.
FT-IR spectroscopy of pollen
Recently Fourier transform infrared (FT-IR) spectroscopy has attracted attention as a method for plant characterisation.1,2,3,4,5,6 As opposed to morphological characterisation of pollen grains with microscopy, infrared spectroscopy offers an operator-independent approach based on chemical characterisation via identifiable spectral features. Roughly, a mid-IR spectrum of pollen contains specific signatures of the constituent biomolecules, such as lipids, proteins, carbohydrates and grain wall biopolymers called sporopollenins (Figure 2). These chemicals are the principal structural and nutritious components of pollen, and they are responsible for the majority of phenotypic attributes. Since the corresponding spectral signals of these chemicals are highly specific, FT-IR spectroscopy is an excellent tool for biochemical analysis of seed plants. FT-IR spectroscopy of pollen can be performed by various sampling techniques, including single reflection attenuated total reflection (SR ATR),1,2,3 transmission measurements in KBr pellets,2,3,4 diffuse reflection (DRIFTS)4 and photoacoustic spectroscopy.5 In addition to these sampling techniques, which are performed by using milligram quantities of a pollen sample, measurements and classifications of single pollen grains can be obtained by using FT-IR microspectroscopy.6 These FT-IR studies have clearly demonstrated that infrared spectra of pollen can be used for simple and rapid plant analyses.
Typical spectra of pollens belonging to conifer and two distinct types of flowering plants [monocot (Carniolan lily) and eudicot (red ash)] are shown in Figure 2, along with vibrational band assignments of their major biochemical components. The two main group of lipids, triglycerides and phospholipids, are characterised by the strong vibrational band at 1745 cm–1 (C=O stretch), as well as by weaker bands at 1462 cm–1 (CH2 deformation) and the 1200–1100 cm–1 spectral region (C–O stretch). In addition, phospholipids have two bands in the 1200–1100 cm–1 spectral region (P=O stretch). Proteins are characterised by two strong and broad bands at 1650 cm–1 (amide I: C=O stretch) and 1550 cm–1 (amide II: NH deformation and C–N stretch). The vibrational bands associated with carbohydrates are predominant in the 1200–900 cm–1 spectral region (C–O–C stretch, C–OH stretch, COH deformation, COC deformation, pyranose and furanose ring vibrations). The sporopollenin bands at 1605, 1515, 1171 and 833 cm–1 can be associated with the vibrations of aromatic rings. Although all pollens are composed of these same chemical building blocks their ratios of biochemicals can vary substantially; this is reflected in their corresponding IR spectra.
, Fraxinus pennsylvanica (Red ash), and Lilium carniolicum (Carniolan lily) pollen. The marked vibrational bands are associated with lipids (L), proteins (P), sporopollenins (S) and carbohydrates (C).")
In general, evolutionarily closely related plants have similar morphological and biochemical traits. This general rule is mirrored in FT-IR spectra of pollen where similar spectral features are shared by evolutionarily related plants. For instance, strong sporopollenin bands, which are dominant in the IR spectrum of Abies cephalonica (Figure 2), are present in the spectra of all pollen species belonging to the pine family (Pinaceae). This is due to a specific physiological feature of pine family pollens. All pine family pollens have a large hollow projection (saccus) from the central body of pollen grain, and two such hemispherical structures can be seen in Figure 1B in Abies cephalonica pollen grain. Since saccus is composed predominantly of sporopollenin, pine family pollens have higher ratio of sporopollenin compared to other biochemicals. On the other hand, pollen grains of cypress family (Cupressaceae) lack saccus features and instead have thick grain walls made of almost pure cellulose. Therefore, pollen grains of these two conifer families can be easily discerned by FT-IR spectra by analysing sporopollenin and carbohydrate bands.2,3
Impact of IR spectroscopic characterisation of pollen on plant biology and ecology
Although, in general, the related pollen species show similar IR spectra, the FT-IR spectroscopic studies have revealed quite significant biochemical differences between closely related species. For instance, DRIFTS and transmission measurements (KBr pellets) were used for the identification of Citrus (citrus genus) pollen species,4 while microspectroscopy was used for identification of Cupressus (cypress genus) pollen.6 These distinctions are rather unexpected since morphologies of pollen belonging to the same plant genus or family seldom display substantial dissimilarities. In practice, due to negligible morphological differences between pollen samples of the related species, pollen grains are rarely identified to the species level. Moreover, identification of the exact plant species is problematic even by means of genetic analysis. We have recently published an FT-IR spectroscopic study of pollen covering more than 300 different plant species. The study revealed huge variations in pollen composition within several plant genera.3 The measured species belonging to Iris (iris), Quercus (oak) and Pinus (pine) genera (as representative for monocots, eudicots and conifers, respectively) show extreme variations in the relative content of triglyceride lipids. It was shown that the difference in the amount of pollen triglycerides can vary ten-fold between two species of the same genus. For example, it has been estimated that pollen of Quercus cerris (Turkey oak) has four times more triglycerides than Quercus robur (pedunculate oak), although both of them were growing at the same location under the same environmental conditions.
Interestingly enough, the same FT-IR spectroscopic study has revealed annual variations in pollen composition as well. In the study, pollen of 15 conifer trees, each belonging to different species of Pinacea family, were monitored during the three consecutive seasons: 2011, 2012 and 2013. The mid-IR spectra showed large inter-annual variations in pollen composition of the measured Pinaceae individuals.
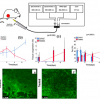
 Mean air temperature for pollen sampling area during 2011, 2012 and 2013 seasons. Pollination onsets of Pinaceae species are indicated with symbols (A: Abies, C: Picea, P: Pinus). (B) Mean air temperature 5, 10 and 20 days prior to the pollination start date. (C) Ratio of absorbance at 1745 cm–1 and 1550 cm–1 in the transmission spectra (KBr pellets) of pollen belonging to Pinaceae samples in 2011 (green), 2012 (red) and 2013 (blue) seasons. Dark and light areas indicate ±1 and ±2 sample standard deviations respectively. Reproduced with permission from Reference 3.")
The variations, predominantly connected to lipid content, are presented in Figure 3 as the ratio of the absorbance of the lipid band (1745 cm–1) to that of the protein band (1550 cm–1). While the pollen composition of the 2011 and 2013 samples is comparable, the composition of 2012 samples deviates significantly. This pattern is in good correlation with temperature profiles prior to the pollination start dates. Figure 3B shows that several weeks prior to pollination, during which the male cones with pollen were developing, the first 11 individuals were under the influence of similar climate patterns: warmer weather in 2011 and 2013, and colder in 2012. The last four pine individuals (the ones that deviate from the other 11 individuals in the inter-annual variations of pollen composition) were under the influence of a different climate pattern.
Traditional biochemical analysis of pollen is done by chemical pre-treatment followed by chromatography or analysis by transmission electron microscopy. Both approaches are time-consuming, expensive and moreover alter the pollen sample in the pre-treatment process. As opposed to traditional methods, FT-IR spectroscopy is a non-destructive analytical technique that enables measurement of samples as found in nature. FT-IR spectroscopy lacks the qualitative precision of the traditional methods that can separate a sample into individual components with methods such as extraction, chromatography and electrophoresis. However, it can provide rapid comparative analysis of a large number of samples with tentative assignment of main groups of biochemicals, such as triglycerides and polysaccharides. As mentioned previously, FT-IR spectroscopy provides information on structural and nutritious components of pollen. Therefore, it offers complementary and often overlooked information to genomic and proteomic methods which measure coding and functional biochemicals, such as nucleic acids and enzymes.
Outlook
The impact of the environment on plant fitness has so far almost exclusively been investigated from the perspective of female functions, while exclusive male traits, such as quality of pollen grains, are mostly overlooked. Recent studies have shown that long-distance pollination is common in plants. Given that single trees can produce millions of pollen grains that could be transported for tens of kilometres, it can be expected that individuals with the fittest pollen grains could rapidly propagate their traits throughout a population. Thus, since it is expected that due to changes in climate different species and populations will have different reproductive success and total plant fitness, methods for monitoring male traits are urgently needed. FT-IR spectroscopy enables simple and rapid identification of pollen by spectral fingerprinting, as well as characterisation and biochemical interpretation related to environment and to taxonomy. Large variations in pollen composition for inter-annual and intra-genus samples, revealed by FT-IR spectra, imply that male functions and pollen competition should be incorporated into ecological and evolutionary studies.
A standardised database of pollen IR spectra, with different taxonomical, temporal and spatial origin, would represent the raw material for a wide range of research, from biology, agronomy and forestry, to ecology. The data should improve comprehension of plant environment interactions, including acclimations and evolutionary adaptations. Therefore, FT-IR spectra could provide valuable information and tools for mitigation and adaptation to global climate change.
References
- E. Gottardini, S. Rossi, F. Cristofoldini and L. Benedetti, “Use of Fourier transform infrared (FT-IR) spectroscopy as a tool for pollen identification”, Aerobiologia 23, 211–219 (2007). doi: http://dx.doi.org/10.1007/s10453-007-9065-z
- B. Zimmermann, “Characterization of pollen by vibrational spectroscopy”, Appl. Spectrosc. 64, 1364–1373 (2010). doi: http://dx.doi.org/10.1366/000370210793561664
- B. Zimmermann and A. Kohler, “Infrared spectroscopy of pollen identifies plant species and genus as well as environmental conditions”, PLOS ONE 9, e95417 (2014). doi: http://dx.doi.org/10.1371/journal.pone.0095417
- C.S. Pappas, P.A. Tarantilis, P.C. Harizanis and M.G. Polissiou, “New method for pollen identification by FT-IR spectroscopy”, Appl. Spectrosc. 57, 23–27 (2003). doi: http://dx.doi.org/10.1366/000370203321165160
- G. Parodi, P. Dickerson and J. Cloud, “Pollen identification by Fourier transform infrared photoacoustic spectroscopy”, Appl. Spectrosc. 67, 342–348 (2013). doi: http://dx.doi.org/10.1366/12-06622
- R. Dell’Anna, P. Lazzeri, M. Frisanco, F. Monti, F. Malvezzi Campeggi, E. Gottardini and M. Bersani, “Pollen discrimination and classification by Fourier transform infrared (FT-IR) microspectroscopy and machine learning”, Anal. Bioanal. Chem. 394, 1443–1452 (2009). doi: http://dx.doi.org/10.1007/s00216-009-2794-